- Research article
- Open access
- Published:
The evolution of the huntingtin-associated protein 40 (HAP40) in conjunction with huntingtin
BMC Evolutionary Biology volume 20, Article number: 162 (2020)
Abstract
Background
The huntingtin-associated protein 40 (HAP40) abundantly interacts with huntingtin (HTT), the protein that is altered in Huntington’s disease (HD). Therefore, we analysed the evolution of HAP40 and its interaction with HTT.
Results
We found that in amniotes HAP40 is encoded by a single-exon gene, whereas in all other organisms it is expressed from multi-exon genes. HAP40 co-occurs with HTT in unikonts, including filastereans such as Capsaspora owczarzaki and the amoebozoan Dictyostelium discoideum, but both proteins are absent from fungi. Outside unikonts, a few species, such as the free-living amoeboflagellate Naegleria gruberi, contain putative HTT and HAP40 orthologs.
Biochemically we show that the interaction between HTT and HAP40 extends to fish, and bioinformatic analyses provide evidence for evolutionary conservation of this interaction. The closest homologue of HAP40 in current protein databases is the family of soluble N-ethylmaleimide-sensitive factor attachment proteins (SNAPs).
Conclusion
Our results indicate that the transition from a multi-exon to a single-exon gene appears to have taken place by retroposition during the divergence of amphibians and amniotes, followed by the loss of the parental multi-exon gene. Furthermore, it appears that the two proteins probably originated at the root of eukaryotes. Conservation of the interaction between HAP40 and HTT and their likely coevolution strongly indicate functional importance of this interaction.
Background
Huntingtin (HTT) is a large intracellular protein with a molecular weight (MW) of 348 kDa, which is functionally involved in diverse cellular processes. These include endocytosis, vesicle transport, autophagy, and transcriptional regulation [1]. A mutation in exon 1 of the HTT gene, which results in the pathogenic expansion of a polyglutamine tract near the N-terminus of the protein, causes Huntington’s disease (HD), a lethal neurodegenerative disease with autosomal dominant inheritance [2].
HTT orthologs are present throughout protists and animals, but absent in plants and fungi [3, 4]. In mice, HTT is essential for embryonic development and viability, since HTT nullizygosity results in early embryonic lethality at about day 8.5 of gestation [5,6,7]. In zebrafish, HTT loss-of-function experiments lead to a variety of phenotypes, including an impact on iron metabolism [8], and different developmental defects, such as impaired neuronal development [9, 10].
Using cryo-electron microscopy (cryo-EM), we recently determined the structure of human HTT [11], which showed a largely alpha-helical protein with three major domains composed of a protein tandem repeat structural motif, the Huntingtin, elongation factor 3 (EF3), protein phosphatase 2A (PP2A), and the yeast kinase TOR1 (HEAT) repeat: a large N-terminal domain with 21 HEAT repeats (N-HEAT), a smaller C-terminal domain with 12 HEAT repeats (C-HEAT), and a connecting bridge domain. Although consisting of homologous repeats, the N-terminal domain forms a one-and-a-half-turn superhelix, whereas the C-terminal domain forms an elliptical ring. Due to the inherent flexibility of HTT [11], structure determination only became possible following the purification of HTT as a tight complex with the cognate huntingtin-associated protein 40 (HAP40). HAP40, formed of 4 canonical and 2 decayed tetratricopeptide repeats (TPR), binds in a cleft between the three domains, forming mainly hydrophobic contacts to N-HEAT and C-HEAT and electrostatic interactions with the bridge domain, thereby stabilizing the conformation of HTT. More specifically, the structure indicated that the C-terminus of HAP40 contains four negatively charged residues (E316, E317, E331, D333), which interact with a positively charged area of the bridge domain. A large number of HTT interactors [1, 12] strongly suggest that HTT serves as a multivalent interaction hub for the coordination of many different functions.
When analysing the interaction of HTT and HAP40 in human 293 cells, HAP40 appeared to be a very abundant interactor of HTT [11]. However, we only obtained the complex between the two proteins upon co-expression, while we could not reconstitute it in vitro from individually purified proteins [11]. The first observation of an abundant, detergent-resistant interaction of HTT with HAP40 in human cell lines was published in 2001 by Peters & Ross [13]. The unusual interaction of HAP40 with HTT, involving coordination of all three HTT domains, explains why in the past HAP40 only rarely surfaced as an interactor of HTT in larger protein-interaction studies unless full-length HTT was used as bait [12, 13]. Interestingly, one of the two studies, which used brain tissue from mice [12], detected HAP40 as the most abundant interactor of HTT, among several hundred less abundant HTT-interacting proteins. These data indicated that the interaction of HTT with HAP40 is not confined to humans.
Very little information on the biological function of HAP40 is available. One research group identified it as an effector of Ras-related protein 5 (Rab5) in endocytosis, mediating the Rab5-dependent recruitment of HTT to early endosomes [14, 15].
In humans, HAP40 is encoded by three sequence-identical paralogs of the factor VIII intronic transcript A (F8A) gene (F8A1, F8A2, F8A3) [16, 17], which all are located on the X chromosome at Xq28. While the F8A1 paralog is contained in intron 22 of the coagulation factor VIII (F8) gene, the other two copies are located outside the F8 gene, closer to the Xq telomere and separated by about 495 kb and 571 kb, respectively, from F8A1. The F8A genes are single exon genes (SEG), i.e. not containing an intron, and are part of a larger, nearly identical repeat sequence of about 10 kb, named int22h-1, int22h-2, and int22h-3.
The F8A genes were first described in the early 1990s [16] when it was noted that recombination between the intra- and extragenic copies of F8A results in haemophilia A due to F8 gene inactivation. This inversion accounts for approximately 50% of all haemophilia A cases [18,19,20].
The extended interaction of HAP40 with HTT, the stability of the complex during purification, and the abundance of this complex both in human cell culture and in mouse brain [12] indicate that HAP40 is an important HTT cofactor. Since no information has been available about the presence of HAP40 in different species, we analysed the evolution of F8A and its potential coevolution with HTT. Substantiating functional importance of the HAP40-HTT interaction, our results strongly suggest the involvement of retroposition, i.e. chromosomal integration of reverse-transcribed mRNA, in the generation of the F8A SEG in amniotes, the coevolution of F8A and HTT, and the conservation of the HTT-HAP40 interaction. Further, our results provide evidence for a homologous origin of HAP40 and N-ethylmaleimide-sensitive factor (NSF) attachment proteins.
Results
Conversion of F8A from a multi-exon to a single-exon gene during the divergence of amphibians and amniotes
While in humans and mice HAP40 has previously been shown to be encoded by single exon genes (SEGs), with three copies present in humans and only one in mice, we noted early on in our study that in zebrafish (Danio rerio) the F8A ortholog (zgc:101679) comprises 11 exons and spans about 9200 nucleotides, versus only about 1700 nucleotides in humans and mice. To analyse the emergence of F8A SEG from an intron-containing ancestor and its possible co-existence with the parental gene in some species, we analysed the genomic organization and chromosomal localization of the F8A locus in 29 representative unikonts (see Additional file 1). Our analyses indicated that F8A is a SEG only in amniotes, whereas it comprises more than one exon in all other analysed species. For instance, a multiple-exon organization of F8A is present in Danio rerio (11 exons), Xenopus laevis (12 exons), and Ciona intestinalis (18 exons). Moreover, a multiple-exon organization of F8A is found in the non-chordates Amphimedon queenslandica (8 exons) and Trichoplax adhaerens (15 exons). In insects, such as Drosophila melanogaster (2 exons) and Bactrocera latifrons (2 exons), a smaller number of exons is annotated.
Copy-number variation and chromosomal location of F8A orthologs
Analysing completely sequenced genomes revealed copy number variation of F8A in different species (see Additional file 1). For example, in the order Primates, humans and orangutans (Pongo abelii) contain three, chimpanzees (Pan troglodytes) and gorillas (Gorilla gorilla) contain two, and gibbons (Nomascus leucogenys) and white-faced capuchins (Cebus capucinus imitator) contain one F8A paralog.
Like in the house mouse (Mus musculus), only one F8A ortholog was detectable in the Norway rat (Rattus norvegicus). In contrast, one or two F8A orthologs are present in laurasiatherians, such as one copy in cats (Felis catus silvestris) and panthers (Panthera pardus), and two copies in horses (Equus caballus), pigs (Sus scrofa), and cattle (Bos taurus). In more deeply-branching species, for example in chicken (Gallus gallus), zebra finch (Taeniopygia guttata), western painted turtle (Chrysemys picta bellii), zebrafish (Danio rerio), pufferfish (Takifugu rubripes), African clawed frog (Xenopus laevis), and tropical clawed frog (Xenopus tropicalis), we identified only one ortholog.
As noted in the introduction, the three F8A paralogs in humans are part of larger repeats (int22h-1, int22h-2, and int22h-3) with a nearly identical sequence. Another SEG, the H2A histone family member B1 gene (H2AFB1), is located in the immediate vicinity of F8A. H2AFB1 codes for an atypical and mammalian-specific histone that is associated with the regulation of apoptosis in spermatogenic cells [21], mRNA processing, and active transcription [22]. When analysing the genomic loci of the F8A orthologs, we observed that the F8A and H2AFB1 genes co-localise in species with more than one F8A gene, such as in human, chimpanzee, orangutan, rhesus monkey, pig, and cattle. In contrast, apart from cats, there is no co-localisation of the F8A and H2AFB1 genes in species with one F8A gene (e.g. in gibbon, mouse, and rat). In cats, the F8A ortholog at locus LOC101095239 is surrounded by two H2AFB1 paralogs (LOC101097798 and LOC101098042), although, according to our analysis, cats possess only one F8A gene.
When determining the chromosomal localization of F8A in 29 representative species (see Additional file 1), we found single-exon F8A orthologs to be almost exclusively located on the X chromosome or in X-chromosome-syntenic regions on other chromosomes. On the X chromosome, single-exon F8A orthologs were always located close to or inside an intron of the coagulation factor VIII (F8) gene. In chicken, F8A is located on chromosome 4 at nucleotides 2,115,165—2,116,487 (GRCg6a), a locus known to be syntenic to the human F8A gene and F8 locus. As the sole exception, in the Norway rat (Rattus norvegicus) the F8A and F8 genes are both located on different autosomal chromosomes, namely chromosome 1 and 4, respectively. Our analysis indicates that the genomic localisation of the multi-exon F8A is not syntenic to the genomic loci of the single-exon F8A orthologs. For instance, the genomic locus of the multi-exon F8A ortholog in zebrafish is on chromosome 1 and not syntenic to the human or chicken F8A loci. Moreover, in some species such as Danio rerio, Ciona intestinalis, Ciona savigny, and Xenopus laevis, the genomic loci of multi-exon F8A orthologs appear to be non-syntenic to multi-exon F8A loci in other species. This diversity in gene structure, copy number, and genomic location suggest a complex history for this family.
HAP40 and HTT are present in all unikonts except fungi
To follow the evolution of HAP40 and HTT, we analysed the non-redundant protein sequence database for their presence either using PSI-BLAST or HHpred. In agreement with published data [23,24,25], HTT orthologs were found in animals (e.g. Amphimedon queenslandica and Trichoplax adhaerens), choanoflagellates (e.g. Salpingoeca rosetta and Monosiga brevicollis), filastereans (e.g. Capsaspora owczarzaki), ichthyosporeans (e.g. Sphaeroforma arctica), and amoebozoans (e.g. Dictyostelium discoideum and Planoprotostelium fungivorum), but not in fungi and nucleariids (Figs. 1 and 2, Additional files 2, 3, 4, 5, 6). We conclude that the common ancestor of unikonts contained HTT and that the protein was lost in the lineage leading to fungi. Outside unikonts, we detected potential HTT homologs in one species each of chromalveolates (the cryptophyte Guillardia theta) and excavates (the free-living amoeboflagellate Naegleria gruberi), but not in archaeplastidans (which include green plants and red algae). Given the very patchy distribution of potential HTT and HAP40 homologs, we cannot judge at present whether HTT and HAP40 originated at the root of eukaryotes, in the Last Eukaryotic Common Ancestor (LECA), and was lost in the plant lineage, or originated in unikonts and was acquired laterally by a small number of other lineages. Strikingly, apart from the parabasalids Trichomonas vaginalis and Tritrichomonas foetus, in which we only detected HAP40, the presence of HTT and HAP40 correlated perfectly across all organisms, supporting the inference that the two proteins evolved together.

Phylogenetic tree of HAP40 (a) and HTT (b). The multiple sequence alignments of HAP40 or HTT orthologs were computed using the MUSCLE algorithm [51, 52] implemented in the MEGA X software [53] as described in the methods section. Phylogenetic trees were calculated by Bayesian inference in MrBayes [55] using Yang’s autocorrelated gamma model [56] and a mixed evolutionary model

adapted from the ‘Tree of Life Web Project’ [69]. Branches with uncertain monophyly are indicated by a dotted line
Phylogenetic distribution of HAP40, HTT, and SNAP. HAP40 and HTT are primarily found in unikonts, whereas SNAP is widespread in all eukaryotic lineages. The tree shown here was
Conservation patterns of HAP40 and HTT, and their interaction
To investigate the conservation patterns of HAP40 and HTT in metazoans, we constructed multiple sequence alignments and mapped their conservation onto the protein structures using the ConSurf server [26]. Alignments of HAP40 from 43 mammals and 73 non-mammals (see Additional file 7) indicated conservation of the N- and C-terminal regions, separated by a variable proline-rich region (41 residues in humans), which is only present in mammals (Fig. 3). Besides, these findings are reflected in the normalized conservation scores computed with the ConSurf method (Fig. 4a, Table 1, Additional files 8, 9, and 12).

Multiple sequence alignment of HAP40 orthologs from representative vertebrates. The alignment, computed using the CLC Main Workbench 7, illustrates the absence of the centrally located proline rich region in non-mammalian species

Evolutionary conservation of HAP40 (a) and HTT (b): Conservation scores were estimated based on a multiple sequence alignment of HAP40 and HTT orthologs from representative species. For the estimation, the ConSurf method [61, 62] with a Bayesian model [4] and the Jones-Taylor-Thornton model [70] was used. Only amino acids that were resolved by Guo et al. [11] (protein database identifier 6EZ8) are shown in the figure
In HTT, the N-HEAT domain (residues 91–1684) with 21 HEAT repeats contains a large insertion between repeats 6 and 7 (residues 400–674), which was unresolved in the cryo-EM structure; the C-HEAT domain (residues 2,092–3,098) with 12 HEAT repeats contains insertions between repeats 1 and 2, and repeats 2 and 3 [11]. The two domains are separated by the bridge domain. The ConSurf conservation scores (Additional files 10, 11, 12) show that the insertion in N-HEAT and insertion 1 in C-HEAT are poorly conserved, insertion 2 in C-HEAT and the bridge domain show intermediate conservation, and the HEAT repeats of N- and C-HEAT are the most conserved parts of the protein (Fig. 4b and Table 1).
In our previous cryo-EM study, we noted that four negatively charged residues in HAP40 (E316, E317, E331, and D333) interact with a positive patch of the HTT bridge domain (K1967, K1968, R1998, R2002, and R2047) [11]. To obtain further information on this interface, we analysed whether mutation of the negatively charged residues of HAP40 inhibits this interaction. We, therefore, performed pull-down assays between wild-type HTT with a poly-glutamine stretch of 17 glutamines (17Q-HTT) and a version of HAP40, in which the four residues were replaced by lysine (HAP40-4 K). Using either HAP40 (Fig. 5) or HTT (data not shown) as bait, our pull-down interaction assays demonstrate an absent or strongly reduced physical interaction between 17Q-HTT and HAP40-4 K fused to a carboxy-terminal or amino-terminal TwinStrep-tag, respectively, in comparison to the wild-type HAP40 (Fig. 5).
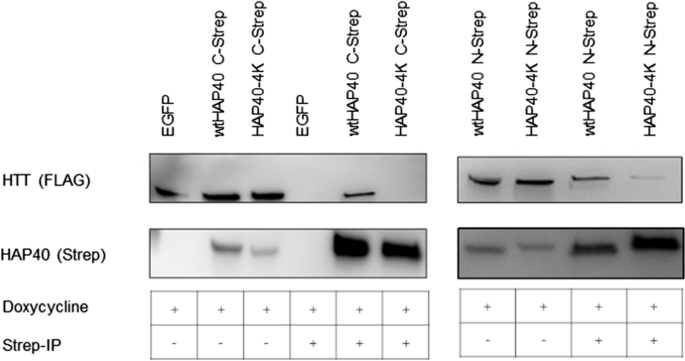
Interaction study of mutated HAP40 and HTT. Human 293-based B1.21 cells, that express 17Q-HTT upon induction, were transfected with plasmids coding for either wtHAP40 or HAP40 mutants in which the E316, E317, E331, and D333 were replaced by lysine (HAP40-4 K). For the interaction study, wtHAP40 and HAP40 fused to a carboxy-terminal and amino-terminal TwinStrep-tag were used. Cell lysates (IP −) or elution (IP +) of the co-immunoprecipitation of wtHAP40 and HAP40-4 K using MagStrep beads (IBA) were analysed by Western-Blot. Western Blots are representative of three independent experiments
Since these data further highlight the importance of the electrostatic interaction between the bridge domain of HTT and the C-terminal part of HAP40, we analysed by bioinformatic methods whether these residues might be evolutionarily conserved. Two interaction patches were suggested by Guo et.al. [11], one formed between the negatively charged E331 and D333 in HAP40 and the positively charged R1998, R2002, and R2047 in HTT, and the second between E316 and E317 in HAP40 and K1967 and K1968 in HTT. In our in-silico analysis, the residues of the first patch were clearly better conserved than average (Table 2) and mostly retained charge complementarity within metazoans, except in insects. In contrast, the residues of the second patch could not be conclusively analysed due to large confidence intervals, but retained charge complementarity in all analysed metazoans, except in Bactrocera latifrons, Amphimedon queenslandica, and Echinococcus multilocularis. We were unable to explore these potential interactions further based on co-evolution analyses (e.g. with complex [27]) due to insufficient depth of the multiple alignments.
To determine whether the physical interaction between HAP40 and HTT is also conserved in deep-branching vertebrate species, we performed pull-down assays between HAP40 and HTT from Danio rerio. To this end, stable HEK293-based cell lines, co-expressing zebrafish HAP40 and HTT, were generated. Using either zebrafish HTT (Fig. 6) or zebrafish HAP40 (data not shown) as bait, our interaction assays demonstrated a physical interaction between zebrafish HAP40 and HTT. Corroborating the conservation of HAP40-HTT interaction in zebrafish further, we could co-purify zebrafish HTT with zebrafish HAP40 from stably transfected HEK293TetOn cells (data not shown).

Interaction study of zebrafish HAP40 and HTT. Zebrafish HTT was immunoprecipitated with magnetic anti-FLAG beads from cell lysates of DrHTT-HAP40 cells. The DrHTT-HAP40 cells constitutively express zebrafish HAP40 and the expression of zebrafish HTT can be induced by addition of doxycycline. Cell lysates (IP −) and eluates (IP +) were analysed by Western-Blot analysis using anti-Strep (HAP40) or anti-FLAG (HTT) antibodies. Western Blots are representative of three independent experiments
Taken together, our bioinformatic analyses and biochemical interaction studies indicate that the physical interaction between HAP40 and HTT is evolutionary conserved at least in vertebrates, but probably throughout animals and, based on the strict co-occurrence of the two proteins, plausibly also throughout unikonts.
HAP40 and NSF attachment proteins are homologous
Since information on the biological functions of HAP40 is limited, we searched for its homologs among proteins of known structure, using profile hidden Markov models. The search was seeded with HAP40 from human, zebrafish, and fruit fly. The best matches, with probability values > 99%, were the mammalian N-ethylmaleimide-sensitive factor attachment proteins α (SNAPA) and γ (SNAPG), and their yeast ortholog Sec17. SNAP proteins regulate vesicle targeting and fusion by orchestrating the interaction between SNAP receptor proteins (SNAREs) and the cytosolic protein N-ethylmaleimide-sensitive factor (NSF) [28,29,30]. The SNAP family is widespread in eukaryotes, with many species comprising multiple paralogs; for instance, while yeast contains one homolog (Sec17), humans contain three homologs (SNAPA, SNAPB, and SNAPG) [31].
In addition to representing the best hits of HAP40 in sequence space, SNAPs are also its best hits by structure comparison (Fig. 7). The best matches, in a search for HAP40-like structures in the RCSB Protein Data Bank using the DALI webserver [32], were to SNAPA, SNAPG, and Sec17, with Z-scores between 16 and 18, and root-mean-square deviations (RMSDs) between 2.5 Å and 3 Å. For comparison, the next best matches are considerably worse, starting at Z-scores of 13 and RMSDs of 5 Å. Like HAP40, SNAPs are composed of 6 TPR hairpins but lack the insertion in repeat 2 and the proline-rich region found between repeats 4 and 5 in mammalian HAP40.

Structure of HAP40 and SNAP proteins. A: Structural alignment of HAP40 (blue, PDB identifier 6EZ8, [11]) with Sec17 (red, PDB identifier 1QQE, [71]), a SNAPA ortholog in Saccharomyces cerevisiae, B: Surface charge distribution of HAP40, SNAPA, and SNAPG from different angles in which a red colouring indicates a negative charge and blue indicates a positive surface charge. Illustration of three-dimensional structure was generated with Chimera X in version 1.13.1 [63]
These comparisons show that HAP40 and SNAPs are each other’s closest relatives in protein databases. This homologous relationship could have resulted from the two families having a common ancestor with 6 TPR hairpins or from their independent amplification to similar structures from an ancestral, single TPR hairpin. We have discussed these two scenarios previously [33]. Global sequence similarity in which TPR hairpin n of one family matches most closely TPR hairpin n’ of the other indicates the former scenario, whereas sequence similarity in which all TPR hairpins of one family match each other more closely than any repeat of the other family indicates the latter. Detailed comparisons of the repeats within and between the two families (Fig. 8) show that for the SNAPs there is a clear signal for amplification from a single TPR hairpin since repeat n matches the other repeats within the protein with high probabilities. For HAP40, there are no corresponding internal matches, each repeat matching only itself with high probability. Instead, outside the self-match, each HAP40 repeat has its best match to the equivalent repeat of SNAP. This shows that both scenarios mentioned above occurred in the evolution of SNAPs and HAP40: an initial amplification gave rise to the SNAP family, one branch of which differentiated strongly to a new form (HAP40), which therefore has its ancestry in a fully formed SNAP-like TPR protein.

Pairwise HMM comparison of HAP40 and SNAP TPR hairpins. The four canonical TPR hairpins of human HAP40, hairpins 1, 3, 4, and 6, and the corresponding hairpins of the yeast SNAP family protein Sec17 were compared to each other using HHpred
Based on the observed sequence and structural similarity, we investigated whether the three human SNAP proteins SNAPA, SNAPB, and SNAPG interact with human HTT. In pull-down assays from cell lysates of HEK293TetOn cells, co-expressing 17Q-HTT together with SNAPA, SNAPB, or SNAPG, we could not detect any interaction using either 17Q-HTT (Fig. 9) or the SNAP proteins as bait (data not shown). The absence of a detectable interaction is not surprising, given that SNAPs display a different surface charges distribution than HAP40 (Fig. 7b).
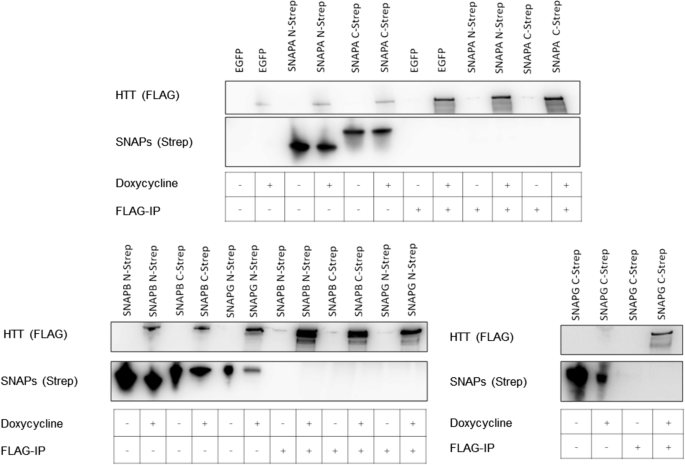
Interaction studies of HTT with SNAPA, SNAPB, or SNAPG using 17Q-HTT as bait. B1.21 cells were induced to express HTT and transfected with pBSK-CMV based plasmids to express SNAPs with carboxy- and amino-terminal TwinStrep-Tags. At 48 h after transfection SNAPs were co-immunoprecipitated using FLAG beads and analysed by western blot. Western Blots are representative of three independent experiments
Discussion
In the human genome, SEGs account for approximately 8.9% of all protein-coding genes [34]. HAP40, a SEG in humans, is expressed from three sequence-identical single-exon paralogs located on the X chromosome, one within and two in the neighbourhood of the F8 gene. Our analysis of the genomic organization of the F8A gene locus across eukaryotes indicated that only amniotes possess the F8A gene as a SEG, suggesting that the conversion from a multi-exon gene (MEG) to a single-exon gene (SEG) took place during or early after the divergence of amphibians and amniotes.
Different mechanisms for the emergence of a SEG from a multi-exon precursor have been discussed. According to one, SEGs arise by duplication events in intron-containing genes [35], in which mostly single exons are duplicated generally leading to truncations [36]. This is not the case in F8A since the SEG and MEG orthologs code for homologous proteins of nearly identical sizes. Another theory posits that SEGs arise by homologous recombination between a reverse transcript of a mRNA intermediate and the genomic locus of the corresponding gene [37]. Our finding that the F8A gene loci in single-exon and multi-exon configurations are not syntenic argues against such a mechanism, although a secondary translocation event cannot be excluded. Retroposition of reverse-transcribed mRNA has been proposed as the predominant mechanism for the generation of functionally active SEGs from parental multi-exon precursors, resulting in retrogenes (also named retrocopies) [34, 38]. Both intron-loss and the observed lack of synteny between the genomic loci of the SEG in amniotes and the MEG F8A gene in zebrafish and other non-amniotic species suggest that retroposition of an intron-less gene copy into a new locus occurred, while the original MEG F8A locus was lost. The lack of a poly-A sequence does not argue against this mechanism, since retroposition in earlier non-mammalian species, such as chicken [39], seems to be mediated by retroviral mechanisms and not by long interspersed elements which would lead to the integration of a poly-A sequence [38].
Strikingly, primates and some laurasiatherians possess more than one F8A paralog, which in humans are contained in a larger int22h repeat. Essentially excluding the possibility that the proteins expressed from the 3 paralogs differ in function, the sequence identity of the three F8A paralogs in humans is maintained by gene conversion [19], i.e. a non-reciprocal transfer of genetic material between paralogs due to homology. Most neutral gene duplications have a low probability to become fixed within a population and are frequently lost or evolve to functionally inactive pseudogenes [40]. The duplicated F8A genes may have been retained during evolution since the duplication event might have conferred higher evolutionary fitness in human primates and laurasiatherians. However, no data supporting the fixation of the F8A duplications have been reported. It is conceivable that for the F8A genes either increases in protein expression levels [41] or subfunctionalisation [42, 43] confer evolutionary advantages; the 16S ribosomal RNA [41] and histone genes [40] are examples for a conferred evolutionary advantage by high expression levels due to gene duplications. On the other hand, subfunctionalisation has been hypothesized as the reason for the fixation of highly similar paralogs [40, 43]. For instance, different paralogs might acquire varying tissue- and development-specific expression profiles, as shown for the engrailed-1 and engrailed-1b [40, 42] paralogs. Since the F8A1 gene is located in intron 22 of the F8 gene in antisense orientation to F8 transcription, HAP40 expression levels may be influenced by transcriptional activity of the F8 gene, a gene that is strongly expressed in hepatocytes. However, as an alternative explanation for amplification and fixation of F8A paralogs in primates and some laurasiatherians, it is also possible that it is rather the very closely linked H2AFB1 histone gene that might have conferred an evolutionary advantage, as it has been observed for other histone genes [40].
At the protein level, human HAP40 contains an internal proline-rich region with a length of 41 amino acids that, according to our data, is absent in non-mammals. In mammals, this region is quite variable with respect to length and amino acid composition. This region had remained unresolved by cryo-EM, indicating flexibility, and was dispensable for the interaction of HAP40 with HTT, as shown by co-expression and protein–protein interaction studies in human cells [11]. Frequently proline-rich motifs are directly involved in protein–protein interactions [44] and the identification of interacting proteins binding to this region will be of significant interest to unravel mammalian-specific functions of HAP40.
HAP40 physically interacts with HTT in cells at significant levels, as shown for human cells cultured in vitro [13], and mouse brain in vivo [12]. In this study, we showed a physical interaction between zebrafish HAP40 and HTT, when expressed in human HEK293TetOn cells. We propose that this interaction is evolutionarily conserved, based on an analysis of the interface observed in our cryo-EM structure. Several charged amino acids predicted in that study to mediate the interaction between the C-terminal region of HAP40 (negative charges) and the bridge domain of HTT (positive charges) are conserved in many orthologs, and their mutation in HAP40 decreased the interaction with HTT.
While HAP40 has been suggested to be an effector of RAB5, information on its biological function has largely remained elusive. The recently determined cryo-EM structure of the HTT-HAP40 complex and its thermal unfolding behaviour compared to the individual proteins alone [11] suggest that HAP40 plays a structural role by coordinating the three domains of HTT, shielding the large exposed hydrophobic surface areas that are distributed over a large part of HTT. Since evolutionary relationships can be employed to infer hypotheses about protein functions, we aimed to identify HAP40 homologs with known functions, which might suggest additional functions for HAP40. In our analysis, HAP40 and SNAPs were found to be the TPR-containing proteins with the highest similarity at sequence and structural levels (Fig. 7), raising the possibility that HAP40 could also be involved in vesicular transport. In line with this reasoning, Pal et al. presented data suggesting that HAP40 mediates the recruitment of HTT and the Ras-related protein 5 (RAB5) to early endosomes [14, 15]. Nonetheless, because TPR-containing proteins exhibit highly diverse functions as scaffold proteins [45, 46], it remains unclear to what extent a potential function of HAP40 can be inferred from its homology to SNAPs, all the more since SNAPs cannot replace HAP40 in its interaction with HTT (this study).
Conclusion
This study is the first analysing the evolution of HAP40, of its encoding gene, the factor VIII intronic transcript A gene (F8A) and the potential coevolution of HAP40 with HTT. HAP40 is encoded by a single-exon gene (SEG) in amniotes, whereas it is expressed from multi-exon genes (MEG) in all other organisms. HAP40 co-occurs with HTT in unikonts, including filastereans such as Capsaspora owczarzaki and the amoebozoan Dictyostelium discoideum, but both proteins are absent from fungi. Outside unikonts, a few species, such as the free-living amoeboflagellate Naegleria gruberi, contain putative HTT and HAP40 orthologs, raising the possibility that the two proteins evolved at the root of eukaryotes.
The interaction between HTT and HAP40, which was shown in humans and mice, also extends to fish, and bioinformatic analyses provide evidence for the evolutionary conservation of this interaction. The closest homolog of HAP40 in current protein databases is the family of soluble N-ethylmaleimide-sensitive factor attachment proteins (SNAPs). SNAPs, however, are unable to replace HAP40 in the interaction with HTT. Taken together, conservation of the interaction between HAP40 and HTT and their likely coevolution strongly indicate functional importance of this interaction.
Methods
Identification of F8A and HTT orthologs in different taxonomic groups
To identify orthologs of F8A and HTT in different species representing the evolution of eukaryotes, sequence comparisons of the human HAP40 (NP_036283.2) and the HTT (NP_002102) reference sequences were performed by discontiguous megablast, PSI-BLAST [47], or HHpred [48, 49]. For PSI-BLAST, the search was performed against the nr70_euk10Jun (2019) database using human HAP40 (UniProt ID: P23610) as query sequence, the BLOSUM60 matrix, an E-value of 1E−3 and an E-value inclusion threshold of 1E−3. HHpred searches were carried out in the MPI Bioinformatics Toolkit against the PDB_mmCIF70 database using default settings, except for the number of target sequences: 1000. Additionally, we performed searches with HAP40 and HTT orthologs, which were identified by initial searches with the human sequences or by database searches in the protein database of NCBI, from Drosophila melanogaster, Amphimedon queenslandica, and Naegleria gruberi. We confirmed all identified hits based on their overall sequence similarity, sequence length, and the e-values resulting from the searches with BALST or HHpred, and by visual inspection of global pairwise sequence alignments against the corresponding query sequences using the Needleman-Wunsch algorithm implemented by the European Bioinformatics Institute (EMBL-EBI) [50].
Calculation of phylogenetic trees
Protein sequences of HAP40 and HTT (Additional file 2) were aligned with the MUSCLE algorithm [51, 52] using the MEGA X software (version 10.0.5, build# 10,180,924-x86_64) [53]. For the MUSCLE algorithm, a gap open penalty of − 2.9 and a gap extend penalty of 0 was used. Moreover, a hydrophobicity multiplier of 1.20 and the unweighted paired-group mean algorithm (UPGMA) with maximal 16 iterations and a minimal diagonal length of 24 was used [51, 52, 54]. Phylogenetic models were calculated by Bayesian inference implemented in MrBayes version 3.2.7.a [55]. For the inference, Yang’s autocorrelated gamma model [56] and a mixed evolutionary model was used. The analysis was conducted for 1,000,000 (HTT) or 500,000 (HAP40) generations on the Baden-Württemberg's high-performance computing cluster (BwHPC). The convergence of phylogenetic trees was determined by the standard deviation of split frequencies which measures the similarity between the tree samples of two independent runs. For HAP40 a standard deviation of split frequencies of 0.011880 and for HTT a standard deviation of split frequencies 0.001884 was reached. Additionally, to test for the robustness of the phylogenetic inference, we computed the phylogenetic trees with the unweighted pair group method with arithmetic mean algorithm (UPGMA) and the maximum parsimony method that are all implemented in the MEGA X software [53] (data not shown).
Analysis of the F8A gene structure and its genomic locus
The HAP40 gene structure was analysed using the National Center for Biotechnology Information (NCBI) Genome Browser. The chromosomal location and the number of exons were assessed in all selected representative species (Additional file 1), which were chosen to represent the main taxa of unikonts. If the F8A gene was not located on the X-chromosome, the F8A gene loci were analysed for synteny using the comparative genomics tools provided by Ensembl (release 97) [57]. Since H2AFTB1 is located in the int22-h repeats in humans and H2AFTB1 orthologs are only described in mammals, we assessed the localisation of F8A in relation to H2AFTB1 in all selected mammalian species.
In some species (Amphimedon queenslandica, Bactrocera latrifrons, Caenorhabditis elegans, Ciona intestinalis, Drosophila melanogaster, Echinococcus multilocularis, Exaiptasia pallida, Schistosoma japonicum, and Trichoplax adhaerens), F8A orthologues were identified by protein sequence, but not at the nucleotide level by discontiguous megablast using the mRNA sequence from Homo sapiens or Xenopus tropicalis. Since some F8A copies might be not annotated, we excluded these species from the analysis of the number of gene copies to avoid the influence of incomplete gene annotation on our results.
Presence of the central proline-rich region of HAP40 in different orthologs
To assess the presence of the central proline-rich region of HAP40 orthologs in all representative species, searches in the protein database of the NCBI were performed with protein–protein Basic Local Alignment Search Tool (BLAST) against the metagenomics protein database provided by the NCBI. The amino acid sequence of human HAP40 (accession number: NP_036283.2) was used as the query sequence. To perform an in-depth analysis of the major taxonomic groups of mammals and non-mammals, we analysed HAP40 sequences from species belonging to common orders of mammals and classes of non-mammals. Therefore, sequences of 43 mammals and 73 non-mammals (see Additional file 7) were analysed by pairwise local sequence alignment with the Smith-Waterman algorithm against the human reference sequence [58] using the pairwise sequence alignment tools of the European Bioinformatics Institute [59] and the Blocks Substitution Matrix 50 [60]. The presence of the central-proline rich region was checked by manual inspection of the resulting alignments.
Estimation of evolutionary conservation with the ConSurf method
Conservation scores were calculated by the ConSurf method [26, 61, 62] based on a multiple sequence alignment of representative HAP40 or HTT orthologs (see Additional files 8, 9, 10 and 11) computed by the MUSCLE algorithm as explained in the section “calculation of phylogenetic trees”. Conservation scores were normalized to a standard deviation of 1 and a mean of 0 [61]. Negative conservation scores indicate higher evolutionary conservation in comparison to other residues of the same protein. The calculated conservation scores were projected onto the HTT-HAP40 protein structure described by Guo et al. [11] (PDB database identifier 6EZ8). Molecular graphics and analyses were performed with UCSF Chimera (version 1.13.1), developed by the Resource for Biocomputing, Visualization, and Informatics at the University of California, San Francisco, with support from NIH P41-GM103311 [63].
Analyses of conservation scores were performed using the R scripting language [64] and the R-packages ggplot2 [65] and readr [66]. The arithmetic means and standard errors of the mean of the conservation score of the amino acid residues in a certain protein domain were calculated (Additional file 12). The positions of the analysed HTT domains were taken from Guo et al. [11].
HMM-based comparison of HAP40 and SNAP TPR hairpins
To investigate the evolutionary origin of the HAP40 and SNAP families, we evaluated the sequence similarity of their TPR hairpins. We chose the four canonical TPR hairpins of human HAP40, hairpins 1, 3, 4, and 6, and their corresponding hairpins in the yeast SNAP family protein Sec17 as representatives. We first searched the nr70 database (NCBI non-redundant protein sequence database clustered at 70% sequence identity) for homologs of human HAP40 and yeast Sec17 using BLAST [47, 67], with E-value threshold (-evalue) set to 0.001 and alignment coverage (-qcov_hsp_perc) to 50%. Separate multiple sequence alignments of HAP40 and Sec17 homologs were parsed out from the obtained hits. These two alignments were subsequently used to extract alignments of the individual TPR hairpins. Profile hidden Markov models (HMMs) were computed from the alignments using hhmake and compared with hhsearch (secondary structure scoring was switched off), both from the HH-suite3 software package for sensitive sequence searching based on HMMs [49].
Interaction studies with mutated human HAP40 and 17QHTT
The HEK293TetOn-based cell line B1.21 [68] was induced for 72 h with 1 µg/ml doxycycline and transfected, using polyethyleneimine, with pBSK-CMV based plasmids coding for wtHAP40 and HAP40-4 K (p.[Glu316Lys;Glu317Lys;Glu331Lys; Asp333Lys]) with either a carboxy- or amino-terminally fused TwinStrep-tag. As a control, B1.21 cells were transfected with a plasmid coding for EGFP. Cells were collected with DPBS followed by centrifugation for 10 min at 400×g. Cell pellets were lysed with 25 mM HEPES, 300 mM NaCl, 0.5% Tween 20, protease inhibitor (Roche), pH 8.0 for 30 min. Lysates were cleared by centrifugation for 10 min at 20,000×g and incubated at 4 °C or 3 h with either MagStrep beads (IBA) or magnetic anti-Flag beads (Sigma Aldrich). Beads were washed three times with 25 mM HEPES, 300 mM NaCl, 0.02% Tween 20, pH 8.0. Bound proteins were eluted with pre-heated SDS loading buffer. Samples were analysed with SDS-PAGE and Western blot analysis using anti-Flag (Sigma Aldrich, 1:2500 dilution) and anti-Strep antibodies (IBA, 1:1000 dilution).
Generation of a stable cell line for co-expression of HTT and HAP40 from zebrafish
A cDNA, human codon-optimized and coding for full-length zebrafish HTT (NCBI NP_571093) and c-terminally fused to a FLAG-His affinity tag, was generated by DNA synthesis (Thermofisher). The cDNA was cloned into plasmid vector pTRE-Tight-BI-AcGFP1 (Clontech) allowing for co-expression of zebrafish-HTT and GFP upon induction with doxycycline (Dox). The resulting plasmid was verified by restriction analysis and transient expression in 293 cells. HEK293 Tet-ON cells (Clontech) were co-transfected with the linearized expression plasmid and a linearized selection plasmid coding for a hygromycin resistance gene. Positive cell clones were isolated by hygromycin selection. A monoclonal cell line expressing zebrafish-HTT (drHTT) was obtained by limited dilution and was validated by Western Blot analysis of cell lysates using a monoclonal anti-FLAG antibody (Sigma Aldrich).
Expression plasmid pBSK/CMV-drHAP40TS was constructed to express, under control of the hCMV promoter, human codon-optimized zebrafish HAP40 (NCBI XP_005160094.1) fused to a c-terminal Twin-Strep-tag. drHTT cells were transfected with pBSK/CMV-drHAP40TS together with a plasmid coding for puromycin resistance. Puromycin-resistant cell clones were isolated and a monoclonal cell line (drHTT-HAP40 3-2-26) was derived by limited dilution. drHTT-HAP40 3-2-26 cells were validated by Western blot analysis of cell lysates with a monoclonal anti-FLAG ab (Sigma Aldrich, 1:2500 dilution) for detection of Dox-inducible drHTT expression, and an anti-Strep antibody (Iba, 1:1000) for detection of constitutive drHAP40 expression.
Interaction studies with HAP40 and HTT orthologs from Danio rerio
The HEK293-based cell line zHTT-zHAP40 were induced for 72 h with 1 µg / ml doxycycline. Not induced zHTT-HAP40 cells were used as control. Pull-down assays were performed as described before.
Interaction studies of SNAPA, SNAPB and SNAPG with human wild-type HTT
Previously described HEK293TetOn-based B1.21 cells, expressing 17QHTT upon induction with doxycycline [68] were transfected with pBSK-CMV-based plasmids expressing human SNAPA, SNAPB, or SNAPG with either carboxy- or amino-terminally fused Twin-Strep tag. Pull-down assays were performed as described before using FLAG beads.
Availability of data and materials
All analysed nucleotide and protein sequences can be retrieved from the nucleotide or protein databases of the National Center for Biotechnology Information (NCBI) with the accession numbers given in the additional files. Moreover, alignments used for the computation of phylogenetic trees and sequence conservation were added to the additional files.
Abbreviations
- F8:
-
Coagulation factor VIII
- F8A:
-
Factor VIII intronic transcript A
- GFP:
-
Green fluorescent protein
- H2AFB1:
-
H2A histone family member B1 gene
- HAP40:
-
Huntingtin-associated protein 40
- HD:
-
Huntington’s disease
- HEAT:
-
Huntingtin, elongation factor 3, protein phosphatase 2A and lipid kinase TOR
- HHpred:
-
Homology detection and structure prediction by HMM-HMM comparison
- HTT:
-
Huntingtin
- LECA:
-
Last Eukaryotic Common Ancestor
- MEG:
-
Multi-exon gene
- NSF:
-
N-ethylmaleimide-sensitive factor
- PSI-BLAST:
-
Position-specific iterative basic local search alignment tool
- RAB5:
-
Ras related-protein 5
- SEG:
-
Single-exon gene
- SNAPA:
-
Soluble N-ethylmaleimide-sensitive factor attachment protein α
- SNAPB:
-
Soluble N-ethylmaleimide-sensitive factor attachment protein β
- SNAPG:
-
Soluble N-ethylmaleimide-sensitive factor attachment protein γ
- SNAPs:
-
Soluble N-ethylmaleimide-sensitive factor attachment proteins
- TPR:
-
Tetratricopeptide-like helical domain
References
Saudou F, Humbert S. The Biology of Huntingtin. Neuron. 2016;89:910–26. https://doi.org/10.1016/j.neuron.2016.02.003.
The Huntington’s Disease Collaborative Research Group. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell. 1993;72:971–83. https://doi.org/10.1016/0092-8674(93)90585-E.
Tartari M, Gissi C, Lo Sardo V, Zuccato C, Picardi E, Pesole G, Cattaneo E. Phylogenetic comparison of huntingtin homologues reveals the appearance of a primitive polyQ in sea urchin. Mol Biol Evol. 2008;25:330–8. https://doi.org/10.1093/molbev/msm258.
Mayrose I, Graur D, Ben-Tal N, Pupko T. Comparison of site-specific rate-inference methods for protein sequences: empirical Bayesian methods are superior. Mol Biol Evol. 2004;21:1781–91. https://doi.org/10.1093/molbev/msh194.
Nasir J, Floresco SB, O’Kusky JR, Diewert VM, Richman JM, Zeisler J, et al. Targeted disruption of the Huntington’s disease gene results in embryonic lethality and behavioral and morphological changes in heterozygotes. Cell. 1995;81:811–23. https://doi.org/10.1016/0092-8674(95)90542-1.
Duyao MP, Auerbach AB, Ryan A, Persichetti F, Barnes GT, McNeil SM, et al. Inactivation of the mouse Huntington’s disease gene homolog Hdh. Science. 1995;269:407–10. https://doi.org/10.1126/science.7618107.
Zeitlin S, Liu JP, Chapman DL, Papaioannou VE, Efstratiadis A. Increased apoptosis and early embryonic lethality in mice nullizygous for the Huntington’s disease gene homologue. Nat Genet. 1995;11:155–63. https://doi.org/10.1038/ng1095-155.
Lumsden AL, Henshall TL, Dayan S, Lardelli MT, Richards RI. Huntingtin-deficient zebrafish exhibit defects in iron utilization and development. Hum Mol Genet. 2007;16:1905–20. https://doi.org/10.1093/hmg/ddm138.
Henshall TL, Tucker B, Lumsden AL, Nornes S, Lardelli MT, Richards RI. Selective neuronal requirement for huntingtin in the developing zebrafish. Hum Mol Genet. 2009;18:4830–42. https://doi.org/10.1093/hmg/ddp455.
Diekmann H, Anichtchik O, Fleming A, Futter M, Goldsmith P, Roach A, Rubinsztein DC. Decreased BDNF levels are a major contributor to the embryonic phenotype of huntingtin knockdown zebrafish. J Neurosci. 2009;29:1343–9. https://doi.org/10.1523/JNEUROSCI.6039-08.2009.
Guo Q, Bin H, Cheng J, Seefelder M, Engler T, Pfeifer G, et al. The cryo-electron microscopy structure of huntingtin. Nature. 2018;555:117–20. https://doi.org/10.1038/nature25502.
Shirasaki DI, Greiner ER, Al-Ramahi I, Gray M, Boontheung P, Geschwind DH, et al. Network organization of the huntingtin proteomic interactome in mammalian brain. Neuron. 2012;75:41–57. https://doi.org/10.1016/j.neuron.2012.05.024.
Peters MF, Ross CA. Isolation of a 40-kDa Huntingtin-associated protein. J Biol Chem. 2001;276:3188–94. https://doi.org/10.1074/jbc.M008099200.
Pal A, Severin F, Höpfner S, Zerial M. Regulation of endosome dynamics by Rab5 and Huntingtin-HAP40 effector complex in physiological versus pathological conditions. Methods Enzymol. 2008;438:239–57. https://doi.org/10.1016/S0076-6879(07)38017-8.
Pal A, Severin F, Lommer B, Shevchenko A, Zerial M. Huntingtin-HAP40 complex is a novel Rab5 effector that regulates early endosome motility and is up-regulated in Huntington’s disease. J Cell Biol. 2006;172:605–18. https://doi.org/10.1083/jcb.200509091.
Levinson B, Kenwrick S, Lakich D, Hammonds G, Gitschier J. A transcribed gene in an intron of the human factor VIII gene. Genomics. 1990;7:1–11. https://doi.org/10.1016/0888-7543(90)90512-S.
Levinson B, Kenwrick S, Gamel P, Fisher K, Gitschier J. Evidence for a third transcript from the human factor VIII gene. Genomics. 1992;14:585–9. https://doi.org/10.1016/S0888-7543(05)80155-7.
Naylor J, Brinke A, Hassock S, Green PM, Giannelli F. Characteristic mRNA abnormality found in half the patients with severe haemophilia A is due to large inversions. Hum Mol Genet. 1993;2:1773–8.
Bagnall RD, Ayres KL, Green PM, Giannelli F. Gene conversion and evolution of Xq28 duplicons involved in recurring inversions causing severe hemophilia A. Genome Res. 2005;15:214–23. https://doi.org/10.1101/gr.2946205.
de Brasi CD, Bowen DJ. Molecular characteristics of the intron 22 homologs of the coagulation factor VIII gene: an update. J Thromb Haemost. 2008;6:1822–4. https://doi.org/10.1111/j.1538-7836.2008.03094.x.
Gu C, Tong Q, Zheng L, Liang Z, Pu J, Mei H, et al. TSEG-1, a novel member of histone H2A variants, participates in spermatogenesis via promoting apoptosis of spermatogenic cells. Genomics. 2010;95:278–89. https://doi.org/10.1016/j.ygeno.2010.02.005.
Tolstorukov MY, Goldman JA, Gilbert C, Ogryzko V, Kingston RE, Park PJ. Histone variant H2A.Bbd is associated with active transcription and mRNA processing in human cells. Mol Cell. 2012;47:596–607. https://doi.org/10.1016/j.molcel.2012.06.011.
Gissi C, Pesole G, Cattaneo E, Tartari M. Huntingtin gene evolution in Chordata and its peculiar features in the ascidian Ciona genus. BMC Genomics. 2006;7:288. https://doi.org/10.1186/1471-2164-7-288.
Myre MA, Lumsden AL, Thompson MN, Wasco W, MacDonald ME, Gusella JF. Deficiency of huntingtin has pleiotropic effects in the social amoeba Dictyostelium discoideum. PLoS Genet. 2011;7:e1002052. https://doi.org/10.1371/journal.pgen.1002052.
Zuccato C, Cattaneo E. Normal function of huntingtin. In: Bates G, Tabrizi S, Jones L, editors. Huntington’s disease. 4th ed. Oxford, New York: Oxford University Press; 2014. p. 243–73.
Ashkenazy H, Abadi S, Martz E, Chay O, Mayrose I, Pupko T, Ben-Tal N. ConSurf 2016: an improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res. 2016;44:W344–50. https://doi.org/10.1093/nar/gkw408.
Hopf TA, Green AG, Schubert B, Mersmann S, Schärfe CPI, Ingraham JB, et al. The EVcouplings Python framework for coevolutionary sequence analysis. Bioinformatics. 2019;35:1582–4. https://doi.org/10.1093/bioinformatics/bty862.
Kaiser CA, Schekman R. Distinct sets of SEC genes govern transport vesicle formation and fusion early in the secretory pathway. Cell. 1990;61:723–33. https://doi.org/10.1016/0092-8674(90)90483-u.
Griff IC, Schekman R, Rothman JE, Kaiser CA. The yeast SEC17 gene product is functionally equivalent to mammalian alpha-SNAP protein. J Biol Chem. 1992;267:12106–15.
Söllner T, Whiteheart SW, Brunner M, Erdjument-Bromage H, Geromanos S, Tempst P, Rothman JE. SNAP receptors implicated in vesicle targeting and fusion. Nature. 1993;362:318–24. https://doi.org/10.1038/362318a0.
Clary DO, Griff IC, Rothman JE. SNAPs, a family of NSF attachment proteins involved in intracellular membrane fusion in animals and yeast. Cell. 1990;61:709–21. https://doi.org/10.1016/0092-8674(90)90482-T.
Holm L. Benchmarking Fold Detection by DaliLite vol 5. Bioinformatics. 2019. https://doi.org/10.1093/bioinformatics/btz536.
Zhu H, Sepulveda E, Hartmann MD, Kogenaru M, Ursinus A, Sulz E, et al. Origin of a folded repeat protein from an intrinsically disordered ancestor. Elife. 2016. https://doi.org/10.7554/eLife.16761.
Jorquera R, Ortiz R, Ossandon F, Cárdenas JP, Sepúlveda R, González C, Holmes DS. SinEx DB: a database for single exon coding sequences in mammalian genomes. Database (Oxford). 2016. https://doi.org/10.1093/database/baw095.
Zhang YE, Vibranovski MD, Krinsky BH, Long M. A cautionary note for retrocopy identification: DNA-based duplication of intron-containing genes significantly contributes to the origination of single exon genes. Bioinformatics. 2011;27:1749–53. https://doi.org/10.1093/bioinformatics/btr280.
Emerson JJ, Kaessmann H, Betrán E, Long M. Extensive gene traffic on the mammalian X chromosome. Science. 2004;303:537–40. https://doi.org/10.1126/science.1090042.
Roy SW, Gilbert W. The evolution of spliceosomal introns: patterns, puzzles and progress. Nat Rev Genet. 2006;7:211–21. https://doi.org/10.1038/nrg1807.
Kaessmann H, Vinckenbosch N, Long M. RNA-based gene duplication: mechanistic and evolutionary insights. Nat Rev Genet. 2009;10:19–31. https://doi.org/10.1038/nrg2487.
International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature. 2004;432:695–716. https://doi.org/10.1038/nature03154.
Zhang J. Evolution by gene duplication: an update. Trends Ecol Evol. 2003;18:292–8. https://doi.org/10.1016/S0169-5347(03)00033-8.
Klappenbach JA, Dunbar JM, Schmidt TM. rRNA operon copy number reflects ecological strategies of bacteria. Appl Environ Microbiol. 2000;66:1328–33. https://doi.org/10.1128/aem.66.4.1328-1333.2000.
Force A, Lynch M, Pickett FB, Amores A, Yan YL, Postlethwait J. Preservation of duplicate genes by complementary, degenerative mutations. Genetics. 1999;151:1531–45.
Lynch M, Force A. The probability of duplicate gene preservation by subfunctionalization. Genetics. 2000;154:459–73.
Williamson MP. The structure and function of proline-rich regions in proteins. Biochem J. 1994;297(Pt 2):249–60. https://doi.org/10.1042/bj2970249.
D’Andrea LD, Regan L. TPR proteins: the versatile helix. Trends Biochem Sci. 2003;28:655–62. https://doi.org/10.1016/j.tibs.2003.10.007.
Perez-Riba A, Itzhaki LS. The tetratricopeptide-repeat motif is a versatile platform that enables diverse modes of molecular recognition. Curr Opin Struct Biol. 2019;54:43–9. https://doi.org/10.1016/j.sbi.2018.12.004.
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–402.
Zimmermann L, Stephens A, Nam S-Z, Rau D, Kübler J, Lozajic M, et al. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J Mol Biol. 2018;430:2237–43. https://doi.org/10.1016/j.jmb.2017.12.007.
Steinegger M, Meier M, Mirdita M, Vöhringer H, Haunsberger SJ, Söding J. HH-suite3 for fast remote homology detection and deep protein annotation. BMC Bioinform. 2019;20:473. https://doi.org/10.1186/s12859-019-3019-7.
Chojnacki S, Cowley A, Lee J, Foix A, Lopez R. Programmatic access to bioinformatics tools from EMBL-EBI update: 2017. Nucleic Acids Res. 2017;45:W550–3. https://doi.org/10.1093/nar/gkx273.
Edgar RC. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics. 2004a;5:113. https://doi.org/10.1186/1471-2105-5-113.
Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004b;32:1792–7. https://doi.org/10.1093/nar/gkh340.
Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018;35:1547–9. https://doi.org/10.1093/molbev/msy096.
Hua G-J, Hung C-L, Lin C-Y, Wu F-C, Chan Y-W, Tang CY. MGUPGMA: a fast UPGMA algorithm with multiple graphics processing units using NCCL. Evol Bioinform Online. 2017;13:1176934317734220. https://doi.org/10.1177/1176934317734220.
Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics. 2001;17:754–5. https://doi.org/10.1093/bioinformatics/17.8.754.
Yang Z. A space-time process model for the evolution of DNA sequences. Genetics. 1995;139:993–1005.
Zerbino DR, Achuthan P, Akanni W, Amode MR, Barrell D, Bhai J, et al. Ensembl 2018. Nucleic Acids Res. 2018;46:D754–61. https://doi.org/10.1093/nar/gkx1098.
Smith TF, Waterman MS. Identification of common molecular subsequences. J Mol Biol. 1981;147:195–7. https://doi.org/10.1016/0022-2836(81)90087-5.
McWilliam H, Li W, Uludag M, Squizzato S, Park YM, Buso N, et al. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013;41:W597-600. https://doi.org/10.1093/nar/gkt376.
Henikoff S, Henikoff JG. Amino acid substitution matrices from protein blocks. Proc Natl Acad Sci USA. 1992;89:10915–9. https://doi.org/10.1073/pnas.89.22.10915.
Glaser F, Pupko T, Paz I, Bell RE, Bechor-Shental D, Martz E, Ben-Tal N. ConSurf: identification of functional regions in proteins by surface-mapping of phylogenetic information. Bioinformatics. 2003;19:163–4. https://doi.org/10.1093/bioinformatics/19.1.163.
Landau M, Mayrose I, Rosenberg Y, Glaser F, Martz E, Pupko T, Ben-Tal N. ConSurf 2005: the projection of evolutionary conservation scores of residues on protein structures. Nucleic Acids Res. 2005;33:W299-302. https://doi.org/10.1093/nar/gki370.
Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin TE. UCSF Chimera—a visualization system for exploratory research and analysis. J Comput Chem. 2004;25:1605–12. https://doi.org/10.1002/jcc.20084.
R Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria; 2020.
Wickham H. ggplot2: Elegant graphics for data analysis. Cham: Springer; 2016.
Wickham H, Hester J, Francois R. readr: Read Rectangular Text Data; 2018.
Camacho C, Coulouris G, Avagyan V, Ma N, Papadopoulos J, Bealer K, Madden TL. BLAST+: architecture and applications. BMC Bioinformatics. 2009;10:421. https://doi.org/10.1186/1471-2105-10-421.
Huang B, Lucas T, Kueppers C, Dong X, Krause M, Bepperling A, et al. Scalable production in human cells and biochemical characterization of full-length normal and mutant huntingtin. PLoS ONE. 2015. https://doi.org/10.1371/journal.pone.0121055.
Maddison DR, Schulz K-S. The Tree of Life Web Project. 2007. https://www.tolweb.org.
Jones DT, Taylor WR, Thornton JM. The rapid generation of mutation data matrices from protein sequences. Comput Appl Biosci. 1992;8:275–82.
Rice LM, Brunger AT. Crystal structure of the vesicular transport protein Sec17: implications for SNAP function in SNARE complex disassembly. Mol Cell. 1999;4:85–95.
Acknowledgements
This work was performed on the computational resource bwUniCluster funded by the Ministry of Science, Research and Arts and the Universities of the State of Baden-Württemberg, Germany, within the framework program BwHPC. We thank Steffen Just for discussion on zebrafish HAP40 and Axel Fürstberger, Johann Kraus and Hans Kestler for initial bioinformatic work.
Funding
Open Access funding enabled and organized by Projekt DEAL. This work was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation – project number 412854449) and by the Max Planck Society. Q.G., W.B. and R.F.-B. have received funding from the European Commission (FP7 GA ERC-2012-SyG_318987-ToPAG). R.F.-B. acknowledges funding from the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) through Germany’s Excellence Strategy—EXC 2067/1- 390729940. The funding agencies provided research support and did not influence study design, collection, analysis, and interpretation of data and writing of the manuscript.
Author information
Authors and Affiliations
Contributions
MS and SK designed biochemical experiments and MS, VA, AL, and SK planned in-silico analysis. TE and BH generated HEK293-based cell lines DrHTT and DrHTT-HAP40. MS and TE performed the co-immunoprecipitation experiment with zebrafish HTT and zebrafish HAP40. In-silico analysis (pairwise sequence alignments, phylogenetic analysis, analysis of protein conservation with ConSurf, and analysis of the f8a gene and its gene locus) were performed by M.S. Structural alignment of Sec17 with HAP40 was performed by MS, QG and SK Co-Immunoprecipitation experiments with SNAPA, SNAPB, SNAPG and 17Q-HTT were performed by MS and TE and experiments with HAP40-4 K were performed by MS, VA performed HMM-based comparison of HAP40 and SNAP TPR hairpins. Data analysis of in-silico and biochemical experiments was performed by MS, VA, AL and SK, MS and VA prepared figures. MS, VA, AL and SK wrote the manuscript and MS, VA, BH, TE, WB, QG, RF-B, AL, and SK revised and commented on the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent to publish
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1.
f8a orthologs in different species. Genes identified by discontiguous megablast using either the f8a from Homo sapiens (NM_012151.3) or Xenopus tropicalis (NM_001078703.1).
Additional file 2.
Accession numbers of identified HAP40 and HTT orthologs.
Additional file 3.
Alignment of HAP40 orthologs used for the phylogenetic analysis.
Additional file 4.
Phylogenetic tree of HAP40 orthologs.
Additional file 5.
Alignment of HTT orthologs used for the phylogenetic analysis.
Additional file 6.
Phylogenetic tree of HTT orthologs.
Additional file 7.
Species used to analyse the presence of the central proline-rich region in HAP40. Protein sequences of HAP40 orthologs from representative mammalian and non-mammalian species were analysed by local pairwise-sequence alignments.
Additional file 8.
Alignment of HAP40 orthologs used for the ConSurf analysis.
Additional file 9.
Raw result of ConSurf analysis for HAP40.
Additional file 10.
Alignment of HTT orthologs used for the ConSurf analysis.
Additional file 11.
Raw result of ConSurf analysis for HTT.
Additional file 12.
R script used for the computation of the domain conservation.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Seefelder, M., Alva, V., Huang, B. et al. The evolution of the huntingtin-associated protein 40 (HAP40) in conjunction with huntingtin. BMC Evol Biol 20, 162 (2020). https://doi.org/10.1186/s12862-020-01705-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12862-020-01705-5